- 移动端


MedChemExpress LLC品牌商
14 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

推荐产品




公司新闻/正文
表观遗传修饰:想要改变基因表达?不只有 DNA,染色体也可以!
94 人阅读发布时间:2026-05-27 12:52
Section.01
表观遗传学
表观遗传学 (Epigenetics) 是指研究通过改变染色体而非 DNA 序列而产生的、可遗传且稳定的基因表达变化的学科。
表观遗传的机制是通过 DNA 碱基的化学修饰和染色体超结构的改变来调节基因表达。表观遗传学的图谱日益复杂,它不仅包含 DNA 甲基化、组蛋白密码、非编码RNA (ncRNA) 和核小体定位,还与 DNA 序列共同构成一个完整的调控体系。表观遗传图谱的紊乱与多种人类疾病的发病机制紧密相关[1]。
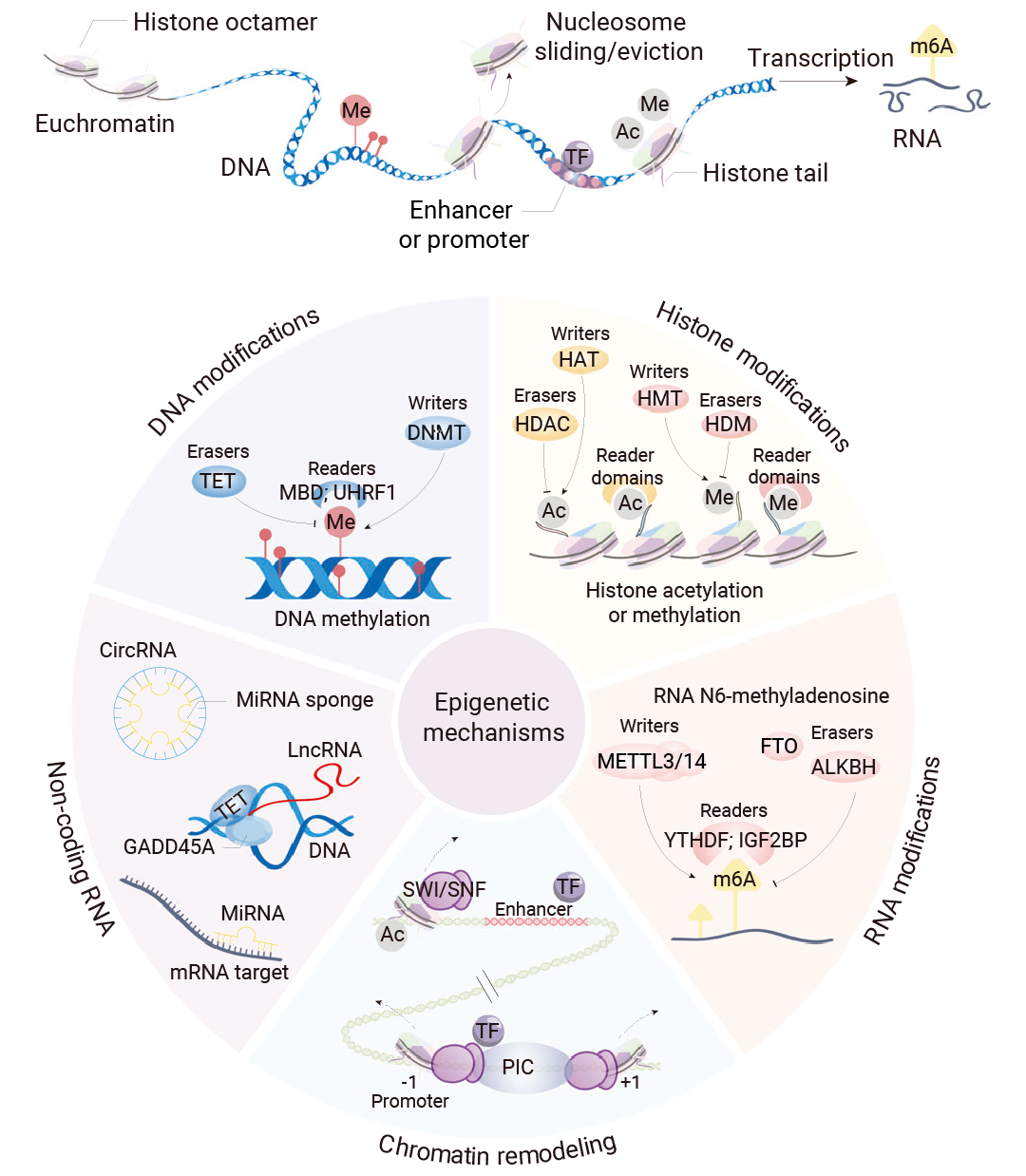
图 1. 表观遗传机制和广泛研究的修饰及其修饰酶[2]。
表观遗传的调控
表观遗传修饰通过三类调控因子实现: (1) 引入各种化学基团的“写入器” (Writers) ; (2) 特异性识别和解读 DNA 或者组蛋白上修饰的蛋白质结构域,即“读取器” (Readers) ; (3) 以及清除修饰的“擦除器” (Erasers) 。
举个例子,表观识别蛋白结构域 (Epigenetic Reader Domains) 是蛋白质上能够特异性识别并结合特定的表观遗传修饰标记的蛋白质结构域,是表观遗传中的“读取器”,将表观遗传标记的化学信息“翻译”成特定的生物学输出,例如基因表达的激活、抑制、DNA 损伤修复或染色质重塑,进而招募其他复合物来影响染色质结构和基因转录[2]。
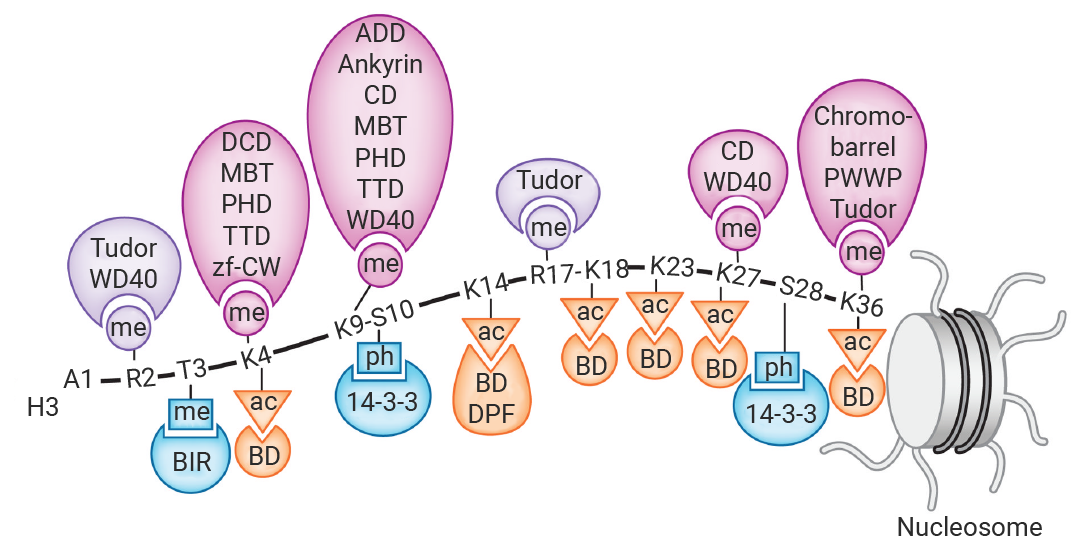
图 2. 组蛋翻译后修饰的“读取器”[3]。
总之,“读取器”、“写入器”和“擦除器”协同作用,协调细胞表观遗传机制和基因表达,并且这些表观遗传修饰因子与多种疾病,尤其是癌症密切相关[2]。
Section.02
DNA 甲基化:
基因的"沉默标签"
DNA 甲基化 (DNA methylation) 是指在 DNA 甲基转移酶 (DNMTs) 的催化下,将甲基添加到 DNA 上的过程。DNA 甲基化发生后,参与基因抑制的蛋白或抑制转录因子被募集与 DNA 结合来抑制基因表达。

DNA 甲基化主要发生在胞嘧啶-鸟嘌呤 (CpG) 二核苷酸位点,尤其是启动子区域内的 CpG 岛。CpG 岛 (CpG Island) 是基因组 DNA 中富含 CpG 的区域,甲基化程度低,在大多数哺乳动物基因组中,CpG 岛位于基因启动子上,使基因就处于“待命”状态。CpG 岛是调控基因表达的“开关”[1][7]。
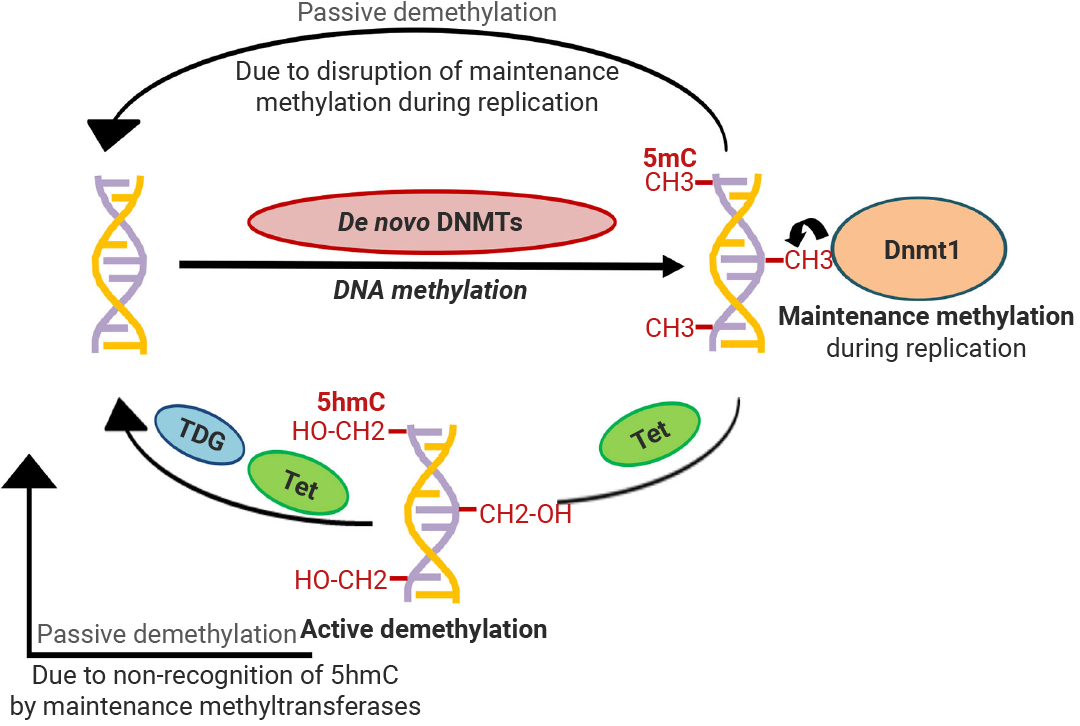
图 3. DNA 甲基化模式[8]。
DNA 去甲基化 (DNA demethylation) 可分为被动去甲基化与主动去甲基化。
被动去甲基化是由于 DNMT1 功能被抑制而引起的,发生在 DNA 复制过程。
主动去甲基化:通过经典通路 TET-TDG-BER 将 5mC 氧化并移除可实现主动去甲基化,不依赖于 DNA 复制。
Section.03
组蛋白修饰:
染色质的"调光开关"
组蛋白 H2A、H2B、H3 和 H4 各两个分子组成八聚体,DNA 缠绕其上形成核小体,并由 H1 组蛋白连接串联。各核心组蛋白均有一条非结构化的氨基末端“尾巴”延伸至核小体外部,为乙酰化、磷酸化和甲基化等翻译后修饰提供位点。
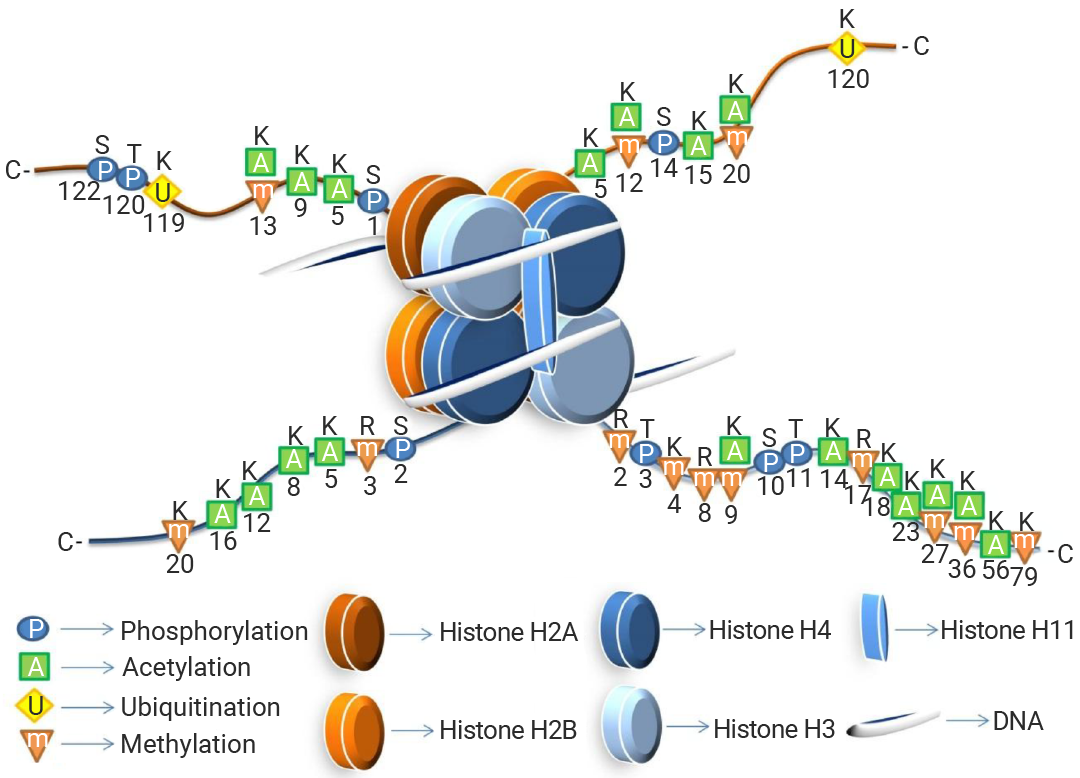
图 4. 组蛋白的结构示意图[9]。
组蛋白修饰的主要方式有 2 种:甲基化和乙酰化。
组蛋白甲基化
组蛋白甲基化 (Histone Methyltransferase) 由组蛋白甲基转移酶 (HMTs) 催化,指在组蛋白 (特别是 H3 和 H4) 的赖氨酸或精氨酸残基上添加一至三个甲基基团的过程。
该过程处于动态平衡,由甲基转移酶 (“写入”) 和去甲基化酶 (“擦除”) 协同调控。组蛋白甲基化的功能复杂,其对基因表达的调控取决于具体位点:例如,H3K4、H3K36、H3K79 的甲基化激活基因转录,而 H3K9、H3K27 的甲基化起抑制作用[10]。
根据 HMTs 催化的氨基酸可分为两大家族:组蛋白赖氨酸甲基转移酶 (KMTs) 和蛋白质精氨酸甲基转移酶 (PRMTs) 。KMTs 家族包括 EZH2、G9a、DOT1L 和 SETD2;PRMTs 家族在哺乳动物中共有 9 个成员 (PRMT1-9)。根据添加的甲基数量,赖氨酸残基上会形成单甲基、二甲基和三甲基化基团 (Kme1、Kme2、Kme3) ,而精氨酸残基上会形成单甲基和二甲基 (Rme1/MMA、Rme2) [10][11]。
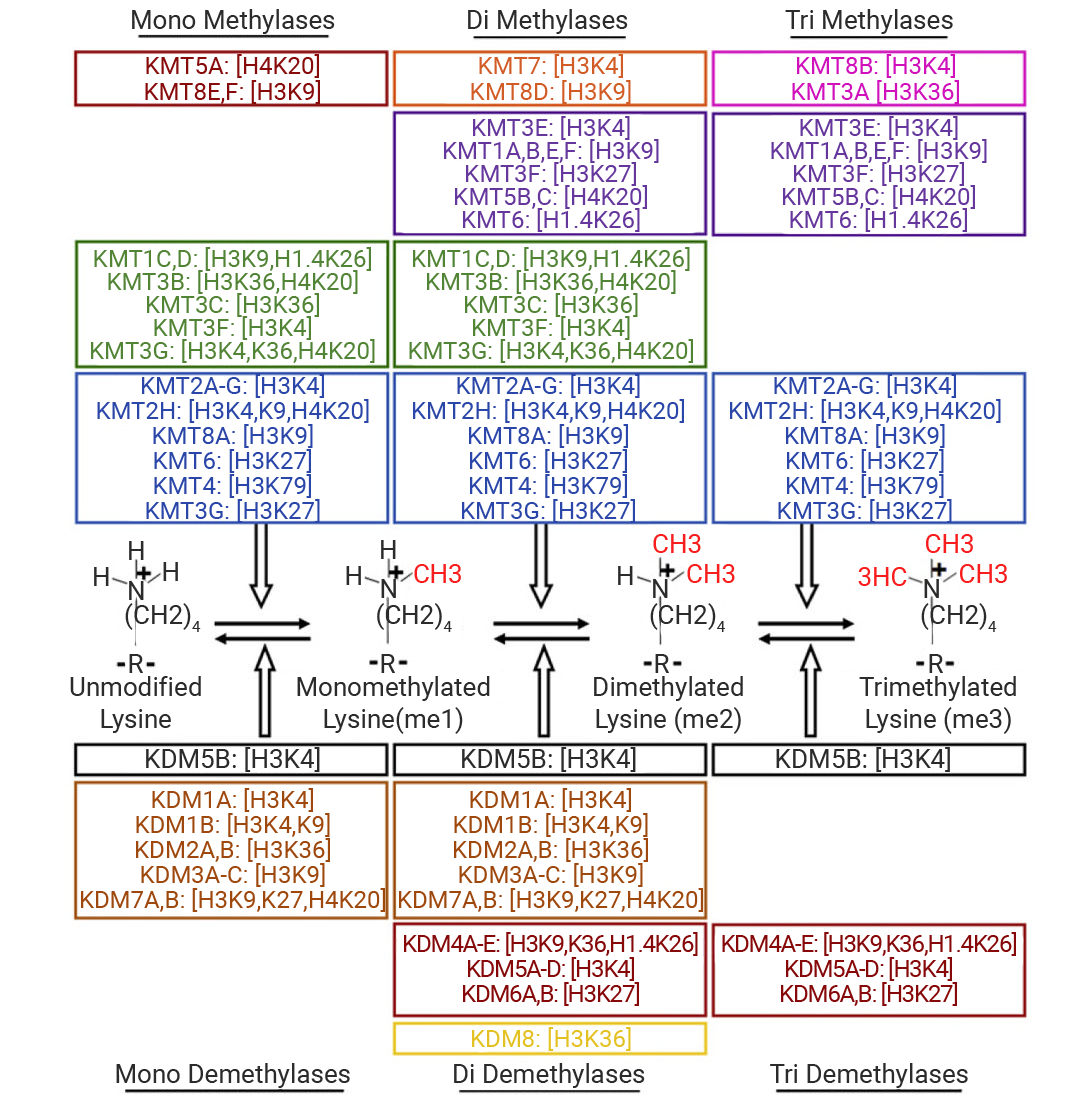
图 5.赖氨酸甲基转移酶 (KMTs) 和赖氨酸去甲基化酶 (KDM) 及其对应底物[11].
几乎所有主要组蛋白赖氨酸甲基化位点的甲基化状态都是可逆的。组蛋白去甲基化由组蛋白去甲基化酶 (HDMs) 催化。HDMs 主要有 LSD 家族 (赖氨酸特异性去甲基化酶) ,如 LSD1 (KDM1A)、LSD2 (KDM1B) 和 JmjC 家族 (Jumonji C 结构域蛋白) ,如 KDM2、KDM3、KDM4 [12] [13] [14] 。
组蛋白乙酰化
组蛋白乙酰化 (Histone Acetylation) 是指在组蛋白 N 端尾部赖氨酸残基上添加乙酰基的修饰,减弱 DNA 与组蛋白之间的相互作用,导致染色质结构从致密变得松散,DNA 更容易被转录因子和 RNA 聚合酶等转录组分接触到,从而激活基因转录。
该过程由组蛋白乙酰转移酶 (HATs) “写入”,组蛋白去乙酰化酶 (HDACs) “擦除”。乙酰化/去乙酰化是一个相对快速的过程,允许细胞迅速响应环境信号 (如激素、应激、营养变化) 来调整基因表达。组蛋白乙酰化水平的异常调控与多种疾病相关,包括炎症性疾病、心血管疾病、神经系统疾病和多种癌症[14]。
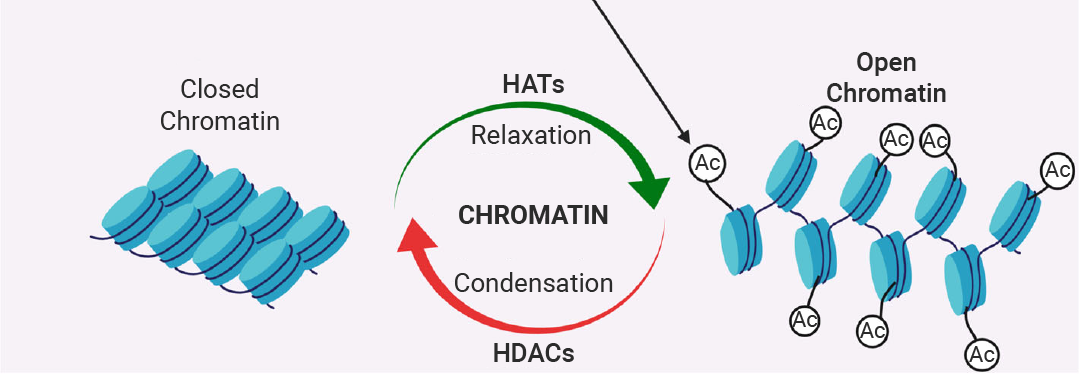
图 6. 染色质受两组酶 HATs 和 HDACs 的调控[14]。
组蛋白去乙酰化酶通过移除组蛋白上的乙酰基团,抑制基因表达并调控染色质结构。在人类中共有 18 种 HDAC 酶,根据其与酵母同源物的相似性和功能差异,分为四个主要类别:
I 类 (Class I):包括 HDAC1、HDAC2、HDAC3 和 HDAC8;
II 类 (Class II):分为 IIa (HDAC4、HDAC5、HDAC7、HDAC9) 和 IIb (HDAC6、HDAC10) ;
III 类 (Class III):也称为 Sirtuins (SIRT1-SIRT7);
IV 类 (Class IV):目前仅包含 HDAC11。
HDACs 的功能异常与多种疾病相关,特别是癌症。在许多肿瘤中,HDAC 活性异常升高,导致抑癌基因被沉默[14]。
Section.04
表观遗传与癌症
癌症是研究最广泛、与表观遗传异常关联最紧密的疾病。癌症表观基因组的特征是 DNA 甲基化和组蛋白修饰模式的整体改变,以及染色质修饰酶表达谱的改变。表观遗传机制的异常在肿瘤发展的各个阶段都发挥着关键作用,包括起始、进展、侵袭、转移以及化疗耐药性的产生。
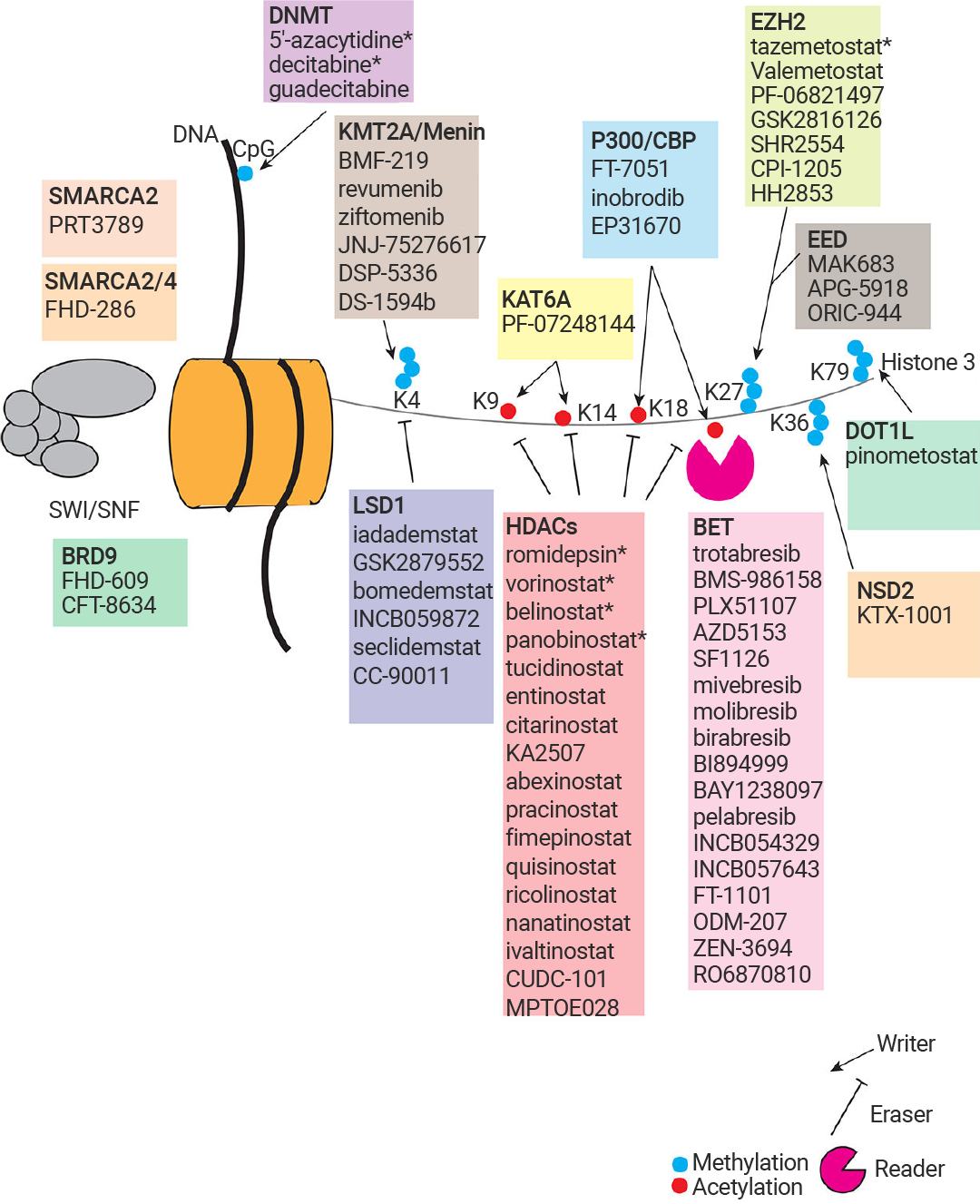
图 7. 癌症中常见的表观遗传靶点[15]。
DNA 甲基化是最早被发现的与肿瘤相关的表观遗传机制。
DNA 低甲基化导致原癌基因的异常激活或基因组不稳定,从而促进癌症的发生和发展。DNA 高甲基化导致肿瘤抑制基因 (如 p16、BRCA1) 和 DNA 修复基因沉默,继而导致这些基因功能障碍,破坏正常的细胞增殖和分化,并促进癌细胞的恶性表型。
DNMT 抑制剂5'-氮胞苷和地西他滨 (Decitabine) 常被超适应症用于治疗急性髓系白血病 (AML) 。组蛋白修饰异常导致致癌基因激活或抑癌基因失活,并且 HDAC 抑制剂已获得 FDA 批准用于治疗皮肤 T 细胞淋巴瘤,如伏立诺他 (Vorinostat) 、罗米地辛 (Romidepsin)。 此外,组蛋白乙酰化修饰的“读取器”溴结构域 (BD) 和末端外结构域 (BET) 家族成员蛋白,包括 BRD2、BRD3、BRD4 和 BRDT,过表达时会促进肿瘤发生。随着表观遗传机制被日益阐明,越来越多的抑制肿瘤的表观遗传靶点药物被开发,并取得显著进展[15]。
靶向表观遗传相关蛋白-蛋白互作的药物发现也有着良好的前景,例如 Menin (MEN1) -KMT2A (赖氨酸甲基转移酶 2A) 抑制剂 Revumenib 已获 FDA 批准用于治疗携带 KMT2A 易位的复发或难治性急性白血病,并于今年 10 月获批用于携带 NPM1 突变的复发性或难治性急性髓系白血病。新一代抑制剂 Ziftomenib、Bleximenib、Enzomenib 等也在临床试验中表现出相似的复合完全缓解率和总缓解率[16]。当然,其他 Menin-KMT2A 互作抑制剂研究成果同样抓人眼球。Jinrong Liang 等科研人员发现,靶向 KMT2A-MEN1 复合物的天然产物臭椿酮 (AIL) 显著抑制了小鼠模型中的骨肉瘤 (OS) 肺转移,KMT2A-MEN1 复合物有望成为骨肉瘤的治疗靶点[17]。
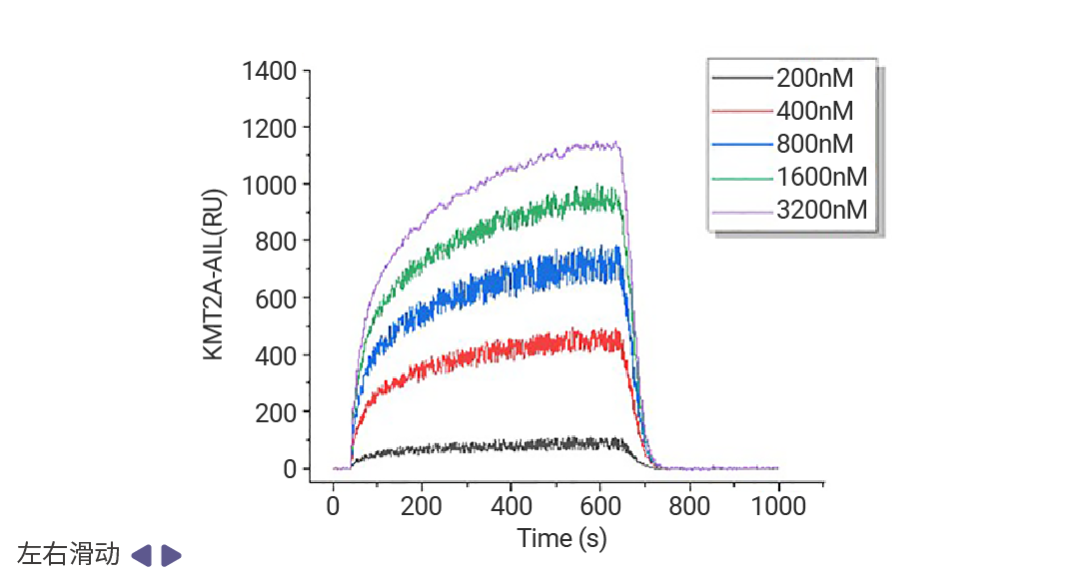
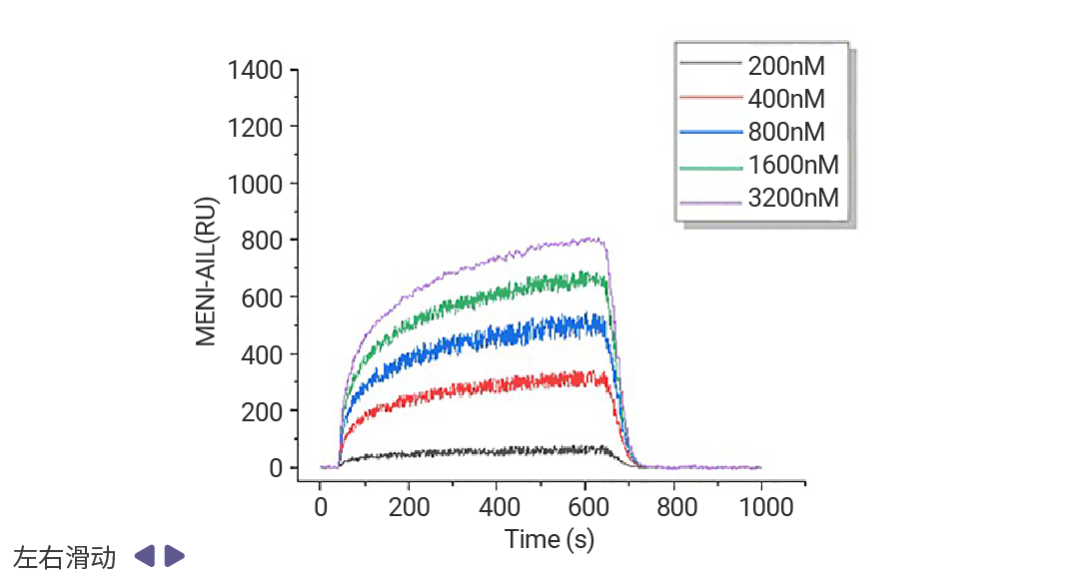
图 8. 表面等离子共振成像 (SPRi) 实验表明,AIL 以剂量依赖的方式与 KMT2A 和 MEN1 结合[17]。
产品推荐
DNMT1蛋白, Human (HY-P703564)
www.medchemexpress.cn/recombinant-proteins/dnmt1-protein-human.html
重组的 DNMT1 蛋白 P26358 (P351-A1600) 片段,由 E. coli 表达,带有 tag Free 标签。
DNMT2蛋白, Human (sf9, GST) (HY-P75711)
www.medchemexpress.cn/recombinant-proteins/dnmt2-protein-human-sf9-gst.html
重组的 DNMT2 蛋白,由 Sf9 insect cells 表达,带有 N-GST 标签。
Histone H1蛋白, Human (His) (HY-P74887)
www.medchemexpress.cn/recombinant-proteins/histone-h1-protein-human-his.html
重组的组蛋白 H1 蛋白,由 E. coli 表达,带有 N-His 标签。
Histone deacetylase 1/HDAC1蛋白, Human (His-SUMO) (HY-P72262)
www.medchemexpress.cn/recombinant-proteins/histone-deacetylase-1-hdac1-human-his-sumo.html
重组的组蛋白脱乙酰酶 HDAC1 蛋白,由 E. coli 表达,带有 N-SUMO, N-6*His 标签。
HDAC6蛋白, Human (His) (HY-P72224)
www.medchemexpress.cn/recombinant-proteins/hdac6-protein-human-his.html
重组的 HDAC6 蛋白,由 E. coli 表达,带有 N-6*His 标签。
Menin蛋白, Human (HY-P702089)
www.medchemexpress.cn/recombinant-proteins/menin-protein-human.html
重组的 Menin 蛋白,由 E. coli 表达,不带标签。Menin 蛋白是一种内分泌肿瘤抑制因子和转录调节因子,也是 KMT2A 蛋白发挥功能所必需的“锚点”,以激活 HOXA 和 MEIS1 等基因。
参考文献
